

-
beat365在线体育官网

-
党建思政
-
科学研究
-
教育教学
-
公开事项
-
校园生活
-
学生工作
PBJ | 湖南农大吴德志团队揭示OsbHLH064调控水稻铁稳态的分子机制
铁元素是植物必需的微量元素,在植物生命代谢过程中发挥关键作用(Briat et al. 2015)。与此同时,铁也是人体必需的微量元素,作物籽粒铁含量直接关系到人类营养与健康。全球约20亿人口受到缺铁性贫血影响,其中以稻米等植物性食物为主食的人群尤为突出。因此,解析水稻铁吸收、转运、存储及其调控机制,不仅有助于增强水稻对不同土壤铁环境的适应性,也为水稻铁营养强化和改善人类营养健康提供理论依据。
近日,Plant Biotechnology Journal杂志在线发表了由beat365在线体育官网/岳麓山实验室吴德志教授团队及其合作团队撰写的“OsbHLH064, an IVb bHLH Transcription Factor, Regulates Iron Homeostasis and Enhances Grain Fe Accumulation in Rice”论文。该研究表明,OsbHLH064缺失突变会导致在铁充足条件下铁稳态相关基因持续激活,而过表达则显著抑制这些基因的表达。此外,OsbHLH064过表达造成水稻地下部、地上部及糙米中铁的过量积累;在铁缺乏条件下,还会诱导活性氧(ROS)过度产生,提示其在铁稳态维持及氧化胁迫平衡调控中具有重要作用。OsbHLH064通过竞争性结合OsPRI1下游靶基因的启动子区域,从而抑制OsPRI1的转录激活活性,进而负调控OsIRO2和OsYSL2等铁吸收和转运相关基因的表达。此外,OsbHLH064不仅能够结合典型的IVb/IVc下游靶基因,还可直接调控一系列参与铁吸收、转运及信号传导过程的关键基因。该研究揭示了OsbHLH064是整合多条铁稳态调控通路的上游核心调控因子,并提示其可作为提高水稻籽粒铁含量的生物强化育种潜在分子靶点(Gao et al. 2026)。

1.OsbHLH064与IVc亚家族bHLH转录因子形成异源二聚体
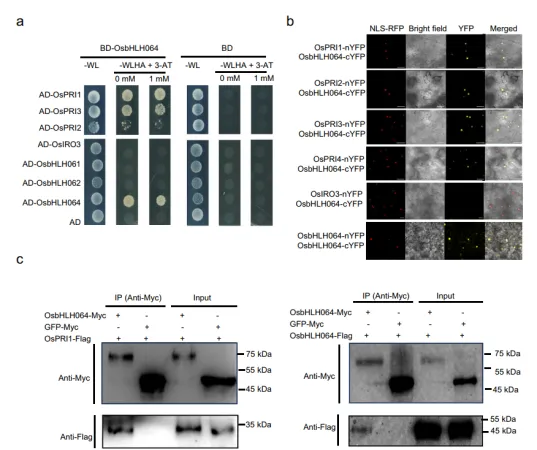
图1.OsbHLH064与IVc bHLH转录因子相互作用
2. IVc bHLH转录因子OsPRIs促进OsbHLH064核积累
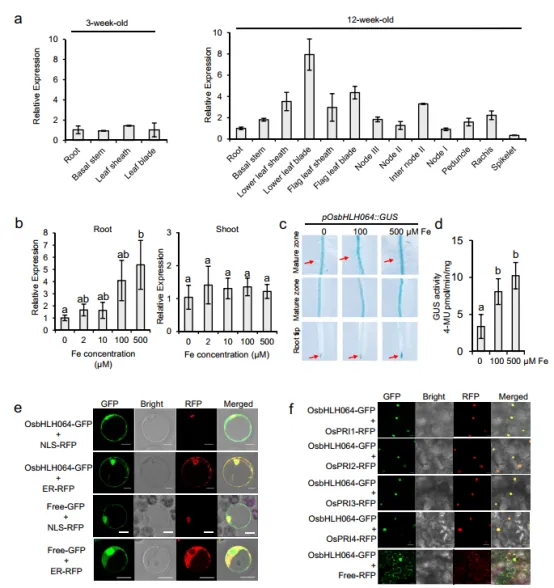
图2.OsbHLH064表达模式和亚细胞定位分析
3. OsbHLH064负调控铁稳态基因表达
4. OsbHLH064过表达诱导非典型缺铁敏感表型及铁稳态紊乱
5. OsbHLH064通过竞争OsPRI1靶基因启动子结合位点调控铁稳态
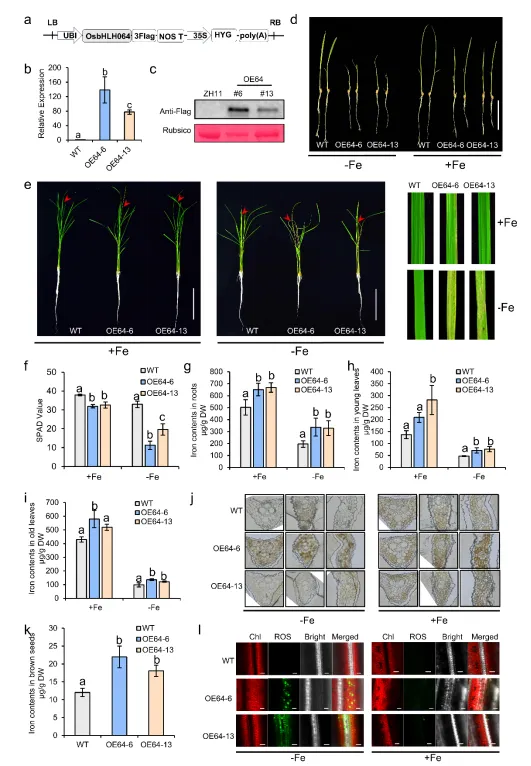
图3. OsbHLH064过表达植株表型分析
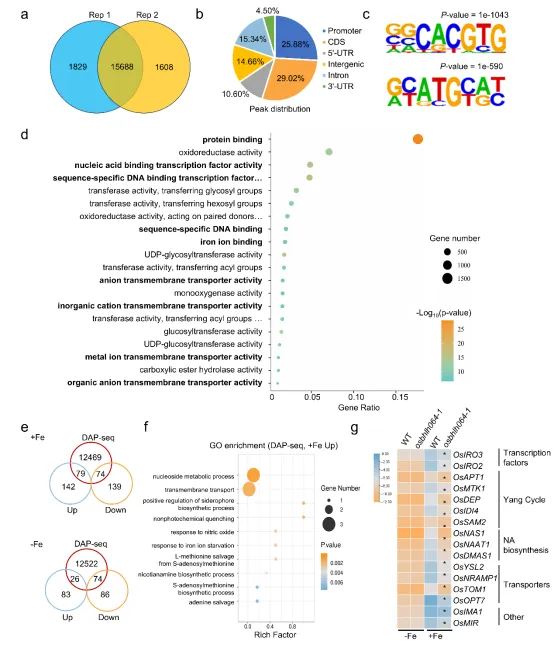
图4. DAP-seq和RNA-seq联合分析鉴定OsbHLH064靶基因
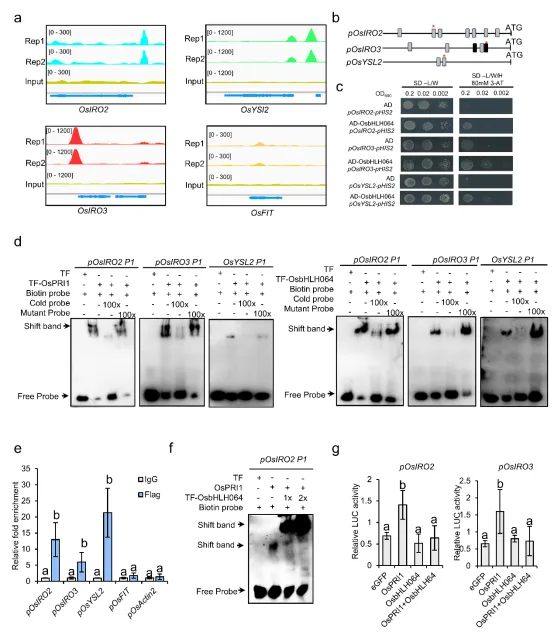
图5.OsbHLH064竞争性结合OsPRI1靶基因启动子
论文链接:
https://onlinelibrary.wiley.com/doi/10.1111/pbi.70593
References:
Briat J-F, Dubos C, Gaymard F (2015) Iron nutrition, biomass production, and plant product quality. Trends Plant Sci 20 (1):33-40
Gao F, Dubos C (2021) Transcriptional integration of plant responses to iron availability. J Exp Bot 72 (6):2056-2070
Gao F, Dubos C (2024) The arabidopsis bHLH transcription factor family. Trends Plant Sci 29 (6):668-680. doi:10.1016/j.tplants.2023.11.022
Gao F, Li M, Dubos C (2024) bHLH121 and clade IVc bHLH transcription factors synergistically function to regulate iron homeostasis in Arabidopsis thaliana. J Exp Bot. doi:10.1093/jxb/erae072
Gao F, Robe K, Bettembourg M, Navarro N, Rofidal V, Santoni V, Gaymard F, Vignols F, Roschzttardtz H, Izquierdo E (2020a) The transcription factor bHLH121 interacts with bHLH105 (ILR3) and its closest homologs to regulate iron homeostasis in Arabidopsis. The Plant Cell 32 (2):508-524
Gao F, Robe K, Dubos C (2020b) Further insights into the role of bHLH121 in the regulation of iron homeostasis in Arabidopsis thaliana. Plant Signaling & Behavior 15 (10):1795582
Gao F, Robe K, Gaymard F, Izquierdo E, Dubos C (2019) The transcriptional control of iron homeostasis in plants: a tale of bHLH transcription factors? Frontiers in plant science 10:6
Gao F, Zhu Z, Xue K, Fu Z, Yue J, Zhang N, Zhou H, Deng Y, Zhou J, Kuang L, Yan T, Li L, Dubos C, Wu D (2026) OsbHLH064, an IVb bHLH Transcription Factor, Regulates Iron Homeostasis and Enhances Grain Fe Accumulation in Rice. Plant Biotechnol J. doi:10.1111/pbi.70593
Liang G (2022) Iron uptake, signaling, and sensing in plants. Plant Communications:100349. doi:https://doi.org/10.1016/j.xplc.2022.100349
Zhang H, Li Y, Pu M, Xu P, Liang G, Yu D (2020) Oryza sativa POSITIVE REGULATOR OF IRON DEFICIENCY RESPONSE 2 (OsPRI2) and OsPRI3 are involved in the maintenance of Fe homeostasis. Plant, Cell Environ 43 (1):261-274
Zhang H, Li Y, Yao X, Liang G, Yu D (2017) Positive regulator of iron homeostasis1, OsPRI1, facilitates iron homeostasis. Plant Physiol 175 (1):543-554
转载网址:https://mp.weixin.qq.com/s/QLoGQ0GRpExY1ECiAjisfQ?scene=1&click_id=1
